-
 大学概要
大学概要
-
 学部・大学院・専攻科
学部
学部・大学院・専攻科
学部- 地域創生学部
- 生物資源科学部
- 保健福祉学部
大学院・専攻科- 総合学術研究科
- 経営管理研究科・HBMS
- 助産学専攻科
-
 学生生活・就職支援
学生生活・就職支援
- 学生生活
- 就職支援
-
 研究・地域連携・国際交流
研究・地域連携・国際交流
-
 入試情報
入試情報
- 学部入試
- 専攻科入試
- 大学院入試
- 出願・合格発表・入学手続
- 大学説明会
- オープンキャンパス
- キャンパス見学
- 進学相談会
- 県大へ行こう-授業公開週間-
- 高大接続・連携事業
- 「受験上の配慮」等について
- 特例措置について
- 入学者選抜試験における個人成績の開示
- 就職・キャリア支援
- 授業料・奨学金・保険
- 科目等履修生・聴講生・研究生
- パンフレット

本文
【生命科学コース・生命システム科学専攻】鞭毛の周期的な屈曲運動を駆動するモーター・ダイニンの活性制御ー八木研究室
八木研究室の論文が Cell structure and function 誌に掲載されました。
IC2 participates in the cooperative
activation of outer arm dynein densely
attached to microtubules
Yusuke Kondo, Tomoka Ogawa, Emiri Kanno, Masafumi Hirono,
Takako Kato-Minoura, Ritsu Kamiya, and Toshiki Yagi
日本語タイトル:
鞭毛の周期的な屈曲運動を駆動するモーター・ダイニンの活性制御
私たちは緑藻クラミドモナスという単細胞の真核生物を用いて,鞭毛運動の研究をしています(図1)。鞭毛あるいは繊毛は細胞表面から伸び出す毛状の運動器官です。それらは,屈曲を根元から先端に向けて周期的に伝搬させて水をかき,細胞本体や細胞外の物質を輸送します。鞭毛・繊毛と異なる名前がついていますが,これらは基本的に同じものですので,ここでは代表して鞭毛として表記します。
鞭毛をその根元,中央,先端,どこで輪切りにしても,「9+2」と呼ばれる共通の構造が現れ(図2),この構造はヒトからゾウリムシまでよく保存されています。外側の微小管には,2つの突起がついていて,そこには,モータータンパク質・ダイニンが含まれています。ダイニンは隣にある微小管を滑らせ,その滑りが鞭毛の屈曲へと変換されます。この鞭毛の屈曲運動は非常に高速です。例えば,クラミドモナスでは1秒間に60回にも達しますが,鞭毛をどうやってこれほど速く動かせるのかまだよくわかっていません。その理由の1つはダイニンの複雑さにあります。ダイニン分子を構成する部品は19種類以上もあり(図3),それぞれの部品の機能はまだ未解明です。鞭毛運動の高速化にはこれらの部品のどれかが特に重要かもしれません。
ここでは,このうちの一つの部品に注目して,鞭毛の高速化機構を調べました。タンパク質の役割を調べる方法の1つに突然変異株の解析があります。クラミドモナスはその遺伝子に変異を入れて鞭毛の運動が異常になった,いわゆる突然変異株を簡単に作れます。この変異した遺伝子と運動性の関係を調べることで,その遺伝子がコードするタンパク質の役割を知ることができます。今回の研究のきっかけは,そのような多数の変異株の中から,見つかった遊泳速度が遅い変異株でした。この変異株はダイニンを構成する部品タンパク質の1つ,IC2のアミノ酸の1つが別のアミノ酸に置換されることで遊泳速度が低下していました。この変異株では,鞭毛が打つ頻度が野生株の70%に低下していることから(図4),IC2が鞭毛運動の高速化に重要であることが初めて分かりました。論文では,この部品タンパク質がどのように鞭毛運動の高速化を行っているのかについて考察しています。興味を持たれた方は論文の方を読んでいただければ幸いです。
この論文では,鞭毛運動におけるIC2というタンパク質の機能を明らかにしました。しかし,鞭毛運動にはまだまだ解明できていない謎が多く,自発的な高速運動機構を理解するためには多くの研究が必要です。八木研究室では鞭毛運動の謎を解き明かすために日々,楽しみながら研究に励んでいます。
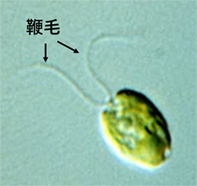
図1 緑藻クラミドモナスの鞭毛
クラミドモナスは鞭毛をヒトが平泳ぎするように動かして泳ぐ。
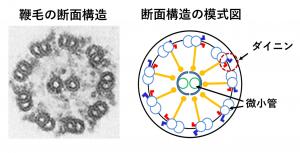
(左)鞭毛横断面の電子顕微鏡写真,(右)その模式図。鞭毛には,微小管が中心に2本,周辺に9本ある。周辺の微小管は2連となっていて,2種類の突起構造(破線の赤と青)がついている。そこには微小管を滑らせるモーター,ダイニン分子が含まれる。
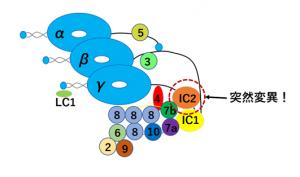
ダイニン分子が多数の部品からなることを示す模式図。本研究は,このうち,IC2(赤い破線)と呼ばれる部品に変異がある場合を調べ,この部品の重要性を調べた。
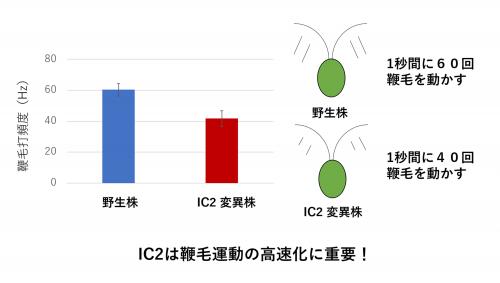
野生株と変異株の鞭毛打頻度(1秒間に鞭毛が何回屈曲するか)を調べた。(左)鞭毛打頻度,(右)それぞれの株の運動を示す模式図。


- 広島県公立大学法人グループサイト


Copyright © 2020 Prefectural University of Hiroshima. All Rights Reserved.