-
 大学概要
大学概要
-
 学部・大学院・専攻科
学部
学部・大学院・専攻科
学部- 地域創生学部
- 生物資源科学部
- 保健福祉学部
大学院・専攻科- 総合学術研究科
- 経営管理研究科・HBMS
- 助産学専攻科
-
 学生生活・就職支援
学生生活・就職支援
- 学生生活
- 就職支援
-
 研究・地域連携・国際交流
研究・地域連携・国際交流
-
 入試情報
入試情報
- 学部入試
- 専攻科入試
- 大学院入試
- 出願・合格発表・入学手続
- 大学説明会
- オープンキャンパス
- キャンパス見学
- 進学相談会
- 県大へ行こう-授業公開週間-
- 高大接続・連携事業
- 「受験上の配慮」等について
- 特例措置について
- 入学者選抜試験における個人成績の開示
- 就職・キャリア支援
- 授業料・奨学金・保険
- 科目等履修生・聴講生・研究生
- パンフレット

本文
【生命科学コース】教員ロングインタビュー4・八木教授
生命科学コースでは、大学Web上で講義、学生のさまざまな活動や卒業生の声を連載形式で紹介しています。本年度は、昨年の新任教員インタビュー(金岡教授)に続き、教員の研究活動やこれまでの歩みについてインタビュー形式の連載としてお届けすることにしました。初回の福永教授、細胞機能制御学研究室の齋藤教授、動物生殖生理学研究室の山下准教授に続き、4回目は八木教授です。
八木俊樹 教授(分子機械学研究室)

分子機械学研究室(3号館7階, 3703号室)
担当講義:物理学II, 分子生物学,生体分子構造学,超分子構造学(大学院)など
Q ご専門や研究内容について教えてください。
私は,細胞の動きに興味をもっていまして,細胞の運動器官,鞭毛・繊毛,が動く仕組みを調べています。鞭毛・繊毛は細胞表面から伸び出す直径が約0.2 μm の毛状器官です。長くて少数の場合を鞭毛と呼び,多数で短い場合を繊毛と呼んでいます。精子の尾部として知られてる鞭毛は(図1A,ウニ精子),根元で作った対称な屈曲を先端に向けて規則正しく伝搬させて波打ち運動を行い,その反作用として細胞は水中を遊泳します。繊毛は,ゾウリムシなどの原生生物の体表によく見られます(図1B)。繊毛の屈曲は片側に強く曲がる非対称なもので,鞭毛に見られる対称な屈曲とは違って見えます。多数の繊毛が協調して非対称な屈曲運動をすると,まるで船の上でオールを漕ぐような要領で,繊毛は細胞本体を動かしたり,細胞外の物質を運搬したりします。後者の例は,私たちの喉の気道に見られます。気道の上皮細胞にある繊毛は,呼気に含まれる細菌やウィルスなどが肺の中に侵入するのを防ぐうえで重要です。気道に侵入した異物は,そこに分泌される粘液に絡め取られて繊毛の運動によって肺側の深部から喉に向かって押し戻され排除されます。ここ2,3年パンデミックを起こした新型コロナウィルスもそのような異物の一つですが,侵入を許すと重い肺炎に発展することはご存知の通りです。
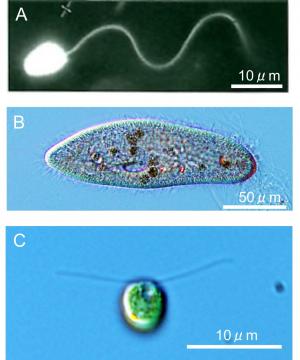
図1.鞭毛と繊毛の例
鞭毛・繊毛は長さも曲がり方も違っていて,一見,異なるもののように見えますが,内部構造は同じで基本的には同じ仕組みで動いていることがわかっています。どちらも輪切りにすると,「9+2」と呼ばれる特徴的な構造が現れます(図2)。どこで輪切りにしても全く同じ構造が現れます(金太郎飴のようです)。つまり,9本と2本の細長いチューブ構造を基本としています。実は,このチューブは細胞内によく見られる微小管と同じものです。周辺にある微小管にはダイニン内腕と外腕と呼ばれる2つの突起がついています。この突起にはモータータンパク質のダイニンが含まれていて,ATPをエネルギー源として隣り合う微小管の間に滑りを起こします。その滑りが鞭毛・繊毛の屈曲運動に発展することが知られていますが,滑りから屈曲への変換機構はよくわかっていません。この「9+2」構造をよく見ていただきますと,微小管とダイニン腕構造だけなく,他にもいろいろな構造があることに気が付きます。「9+2」構造の中には全部で300種類以上の部品タンパク質が含まれていて,それらは,微小管の滑りを調節して複雑な屈曲を生み出すうえで,それぞれが固有の役割を持つと考えられています。それらの役割がわかれば,波動運動の仕組みが理解できそうです。でも,300種類もある部品をひとつひとつ取り出して調べるのは大変です。逆に,たくさんある部品のうち特定の部品だけを壊すことができれば,壊したあとで鞭毛・繊毛の動きがどう変わるかを調べれば,それぞれの役割がわかりそうです。そのような実験は,突然変異株を利用すれば可能です。
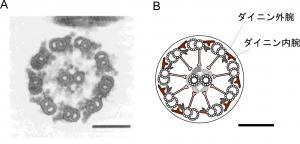
図2.鞭毛・繊毛の横断面
A.電子顕微鏡写真, B.横断面模式図 バーは0.1 μm。
私は,緑藻クラミドモナス(図1C,単細胞で鞭毛を2本もっています)の突然変異株を使って,鞭毛の運動に重要な部品タンパク質を調べています。クラミドモナスに紫外線を当てると,遺伝子DNAにランダムに変異が入ります(配列が書き変わったり,一部の配列が無くなったりします)。そうなると,その遺伝子がコードするタンパク質が異常になり,鞭毛の運動が変化した突然変異株がたくさん取れてきます。私はこれまでに,そのような変異株を取ってきては,どの遺伝子がおかしくなったか,どのタンパク質に異常があるか,また,そのタンパク質は鞭毛内のどこにあり,鞭毛運動においてどんな役割をしているのか,を調べてきました。例えば,さきほどお話しした,モータータンパク質のダイニンには実は15種類もあること,その中には,べん毛の根元から先端まで均一に存在するものもあれば,根元にだけ存在する変わったダイニンもあることがわかってきました(図3)。今,それらのダイニンを失った変異株を使って,鞭毛運動における各ダイニンの役割の違いを調べているところです。
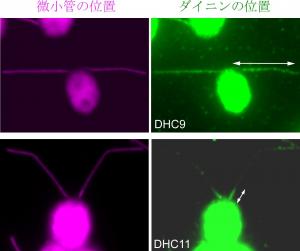
図3.鞭毛根元にだけ存在するダイニン
多種類あるダイニンごとに,鞭毛上に並ぶ位置が異なっていた(上)鞭毛上に一様に存在する場合,(下)鞭毛根元にだけ存在する場合。 特異的な抗体でそれぞれの場所を特定した実験。
Q 大学生の頃はどう過ごされていましたか?
今でこそ生命科学コースの教員ですが,実は高校生の頃は生物学が嫌いでした。暗記が中心だった高校生物について行けないなと思ったことが原因です。一方,物理学と化学は,ある現象はこう考えると説明できますとか,こういう法則があるからこの反応は次にこうなると予測できます,というようなことで私にとってはわかりやすく感じました。それで,2年生からはこの2科目を選択して,大学では理工学部の物理学科に進むことにしました
大学ではごく普通の理系学生でした。講義の合間に友達に誘われて雀荘に行ったり,近くの3本立ての安い映画館で時間をつぶしたりしながら,結構多かった実験実習とそのレポート作成,それと演習(ひたすら問題を解く)に追われていました。講義はそれぞれ新鮮で面白く,結構まじめに聞いた気がします。そんな中,中高の教員免許を取る都合上,どうしても生物学の講義を取る必要があって仕方なく講義を受講したのですが,これが意外にも面白い。高校の生物学とはまるで別物のように感じられたのです。たぶん,担当した先生が,物理学科の学生でも興味を持つようにずいぶん噛み砕いてアレンジされたのだと思います。生物を見る見方にはいろいろあることに気づかされ,私にとっての転機になった講義でした。
4年生になると,研究室に配属されてそれぞれの研究テーマを深く「研究」する卒研が始まります。物理学の花形は,素粒子物理学,宇宙物理学,物性物理学等ですが,私がいた大学には,なぜか「生物物理学」の研究室が3つもありました(私の大学は,生物物理学研究を標榜する日本におけるメッカの1つだったのですが,恥ずかしながら,私は知らなかったのです)。3年生の講義で生物学に対する認識を変えていた私は,生物物理は新しくて面白そうと思って,この3研究室に狙いを定めます。「生」の試料を扱うほうが具体的で面白そうだと思ったので,3研究室の中から,「筋肉」の研究室を志望して無事配属されました(他の2つは,タンパク質立体構造の物理学,視覚の物理学で,どちらも理論的な研究でした)。生物の知識がほぼゼロの状態でしたので,最初は大変でしたが(ピペットの使い方さえ怪しい),ひよこのように先輩にくっ付いて修行するうちに何とかサマになっていった気がします。毎日,終電近くまで研究室にいましたが,日々新鮮で苦にはなりません。そのうち,自分の研究テーマが決まります。「筋肉の収縮を顕微鏡下で再現する」という何ともわかりやすいものでした。具体的には,モータータンパク質・ミオシンを固定したガラス上をアクチンフィラメントがATP存在下にするする滑る様子を見る,というものです。試行錯誤してアクチンが滑る様子がようやく見えたときはとてもうれしかったことをよく覚えています(そのため,視力が大分落ちましたが)。この実験は,アクチンフィラメントが滑るだけでなく回転運動もするという新発見にもつながりました。実験は楽しかったのですが,実験材料の調整(ウサギから筋肉をとってくる)には結構抵抗があり,そこが,その後の進路選択に影響しました。
Q 研究者になろうと思ったきっかけは何ですか?
特に分野は決めていなかったのですが,小学生の頃から研究者になりたいなと思っていました。子供の頃,私の兄がなぜか稲に興味をもっていまして,その兄が奈良県の農業試験場(奈良の出身です)に見学に行く機会があり,それに私もついて行ったのが研究を意識し始めたきっかけだった気がします。そんな兄は文系に進み,今は普通の会社員ですので,人生どうなるかわかりません。 さて,大学4年生になった私は,理系に来たのだから大学院に進みたい,せっかく始めたのだから生物が研究できるところがいいなと思い始めまして,所属大学以外も視野に入れて,行先を探し始めました。ところが困ったことに,生物の研究ができる大学院で物理学を受験科目に入れているところがほとんど見当たりません。生物学の知識がほぼない私には,生物学での受験は無理そうです。さてどうするか? 配属先の先生(石渡信一先生)に相談したところ,先生ご自身の出身研究室を薦めていただきました(そこは,いわゆる大学院大学で,生物物理学の研究室は物理,化学,生物どれでも受験可でした)。薦められるままにそこを受験して,(名古屋にある)その研究室に進むことにしました。研究室を変えた理由はいくつかありましたが,その一つだったのが扱う研究材料です。新しい研究室では,今でも研究を続けているクラミドモナスを使って研究をしていました。それはウサギと違って,研究でいくらたくさん使ってもあまり罪悪感がありません。私にとってそれは大きかったのです。
新しい研究室(神谷律先生)に移って,クラミドモナスの鞭毛運動の研究を始めます。材料が変わっても研究方法は本質的にあまり変わりません。それは当たり前で,筋肉のアクチン・ミオシン,鞭毛のダイニン・微小管は,どちらもATPをエネルギー源として滑り運動を行う点で同じです。そのため,あっという間に新しい研究環境に慣れ,鞭毛の動きを細かく調べる装置(ナノメートルレベルの動きを調べることが可能)を作って,「9+2」構造を構成する部品の役割を調べました。いろいろ結果が出て来ると研究は面白くなるもので,大学の同級生がほとんどが迷わず就職していく中(世間はバブル真っ只中で,就職先に困っている人はいない状態),博士課程に進むことにしました。自分自身,とにかく楽観的でまあ何とかなるだろうと思っていたわけですが,所属した研究室では博士課程に進むのが当たり前という雰囲気があったことも影響したと思います。
Q 広島に来られたきっかけは何ですか?
博士課程を修了して,2年間は日本学術振興会の研究員に採用されて(前回の山下先生の紹介記事を参照)そのまま東京の神谷先生の研究室で研究を続け,つづいて,運よく,同じ研究室で助手(のちに助教)に採用されました。10年近くその職にとどまった後,京都,東京と,関西・関東を行ったり来たりした後,2014年に縁あって,ここ県立広島大学に職を得ました(広島に来て,今年で10年目に突入です)。偶然にも,私の前の研究室の教授(相沢慎一先生)は神谷先生の後輩です。神谷先生は石渡信一先生(私が卒業した大学研究室の教授)の後輩です。私自身,(源流をたどると)各先生方の出身研究室につながります。気が付いてみるとすごい縁だなと感じるところです。 これまで学会などで広島を通過することはありましたが,住むのは初めてです。こちらに来るまで,原爆ドーム,安芸の宮島には行ったことがありませんでしたが,ようやく研究室内の旅行で行く機会がありました(図4)。それぞれに印象深いものがありますね。今後は,瀬戸内の島々などにも行ってみたいと思っています。

図4. 留学生と研究室メンバーで厳島神社へ
Q 趣味などはありますか?
特に今は何もやっていませんが,東京では一時期,馬に乗っていたことがあります。多摩川のほとりの乗馬クラブに通っていたのですが,希望者は馬に乗ったまま多摩川の浅瀬をわたり対岸に渡るなどというイベントがあり,それに参加したことがあります。真冬の早朝に川面から湯気がたつ中を,馬と一緒に闊歩できたのはなかなか貴重な体験でした。忙しくなってやめてしまってずいぶん経ちます。ここ庄原では乗馬ができるところもあるようですので,機会があれば再開してみたいと思っています。その他と言えば,本を読むことが好きで,専門書だけでなく一般書も置いている大学の図書館には,ちょくちょく顔を出しています。
Q 最後に高校生や在学生へのメッセージを
生物資源科学部のある庄原市は,広島県を長方形とすると,ちょうど広島市と対角線で結んだそれぞれ頂点の位置にあります。広島市内から離れていて確かに田舎ですが,良いところもたくさんあると思います。私がこのキャンパスを見ての第一印象は,アメリカの大学に似ているなというものでした。留学したことのある米国マサチューセッツ州の大学(UMASS Medical School)は内陸の結構な田舎にありました(州の第2都市のウースター,第1の都市はボストン)。湖のほとりに建つ丘の上の大学という点で地理条件もよく似ています。他にも,米国の歴史ある優秀な大学は結構な田舎にあることが多いです(Ivy league の ひとつとして知られるCornell 大学なども)。インターネットで世界と結ばれている昨今,田舎にいてもそれほど世間と隔絶されることもない気がしますし,かえって大都市よりもごちゃごちゃしていなくて落ち着くように思います。人生の一時期,そういった環境に身を置くのもいいかなと思いませんか?
若い皆さんは(高校生も大学生も)特にこれと決めずにいろいろな勉強,体験をするのがよいと思います。こんなこと,将来役に立つの?と思うこともあるかもしれません。私もそう思いながらいろいろやっていた時期がありますが,振り返ると,それぞれの経験は何かの形で自分の身になっているように思います。ぜひ,いろいろなことに挑戦してください。私たち教員はいつでも皆さんの手助けをします。困ったら声をかけてください。
研究室および教員に関する情報 *より詳しい教育・研究内容は以下をご覧ください。
卒業生の声:


- 広島県公立大学法人グループサイト


Copyright © 2020 Prefectural University of Hiroshima. All Rights Reserved.